Page 2 - REMERCIEMENTS
REMERCIEMENTS S’il est une chose merveilleuse que j’ai apprise durant ces années de thèse, c’est ta présence en moi. J’ai appris à réaliser que tu es toujours là, dans cette place secrète qui est en moi, que tu me vois, tu m’entends, et tu exauces toujours mes prières ; que tu me guides dans mes cho...
Page 4 - Résumé; Daphnia pulex
Résumé Résumé Face à l’urgence exprimée par la communauté internationale de trouver des solutions efficaces et durables à la crise sanitaire, due au manque d’accès à l’assainissement dans les pays du sud, la présente thèse s’est intéressée à l’étude de la possibilité de promouvoir ce secteur à trave...
Page 5 - Les résultats des analyses de stœchiométrie révèlent que :
Résumé Les résultats des analyses de stœchiométrie révèlent que : - Pour produire 1 g d’équivalent DCO de D. pulex, 5 g d’équivalent DCO de Scenedesmus sp., (ou d’ E. coli) sont oxydés dont 0,77 sont convertis en matières organiques particulaires et, le reste (soit 3,23 équivalent DCO ou 65%) est ox...
Page 7 - Publications réalisées dans le cadre de la présente thèse; pulex
Publications réalisées dans le cadre de la présente thèse Liady M. N. D., Fiogbe E. D., Cauchie H. M., Vasel J. L. 2013. About the interest of a zooplankton compartment in pond systems: Methodology to study the growth of D. pulex . Oral communication in the 10th International Water Association Speci...
Page 8 - Table des matières; Chapitre II: COMPTAGE ET ESTIMATION DE BIOMASSE DE
Table des matières INTRODUCTION GENERALE………………………………………………………………………………………………………..- 1 - I. Contexte et justification ................................................................................................. - 1 - I.1 Le contexte strategique mondial ...............................................
Page 9 - Scenedesmus
III.4 Relation poids sec - taille des individus .............................................................. - 61 - IV. Conclusion ................................................................................................................ - 65 - V. Références ....................................
Page 12 - Liste des figures; et al
Liste des figures Liste des figures Figure I.1: Schéma de fonctionnement d’un bassin facultatif (Tchobanoglous et al ., 1985 ; cités par Shilton, 2001) Figure I.2 : Schéma synthétique de description des différents bassins de lagunage Figure I.3 : Vue microscopique de quelques protozoaires (Leclercq ...
Page 13 - Scenedesmus sp
Liste des figures Figure II.15 : Corrélation entre Axe majeur mesuré au microscope et Axe majeur estimé par le modèle Figure II.16 : Test de normalité de la distribution des biais relatifs (%) Figure II.17 : Relation entre poids sec et Axe majeur de D. pulex Figure II.18 : Relation entre poids sec e...
Page 15 - M aeruginosa
Liste des figures Figure IV.24 : Evolution des moyennes de teneurs en oxygène dans les cultures de D. pulex sur Scenedesmus sp . Figure IV.25 : Evolution des pH moyens dans les cultures de D. pulex sur Scenedesmus sp . Figure IV.26 : Estimation du taux spécifique de croissance d' E. coli sur le mili...
Page 17 - Liste des tableaux
Liste des tableaux Liste des tableaux Tableau I.1: Quelques modèles empiriques de dimensionnement du bassin facultatif Tableau I.2: Conception et dimensionnement des principaux types de stations d’épuration par lagunage à travers le monde (d’après Vasel et VanderBorght, 1998 ; Mara, 1995; Mendes et ...
Page 19 - acuminatus
Liste des tableaux Tableau III.17: Analyse de significativité du biais relatif moyen par rapport à 0, des estimations de la densité cellulaire de M . aeruginosa par spectrophotométrie Tableau III.18: Test du modèle complet MES M. aeruginosa = f(Abs 760nm ) Tableau III.19:Paramètres estimés du modèle...
Page 22 - réaction traduisant le processus de respiration
Liste des tableaux Tableau VI.18 : Vérification des bilans des atomes, des charges et de la DCO de l’équation de réaction traduisant le processus de respiration Tableau VI.19: Matrice partielle de Petersen relative à la respiration de D. pulex Tableau VI.20: Valeurs des paramètres stœchiométriques u...
Page 24 - Introduction générale; INTRODUCTION GENERALE; I.1 Le contexte strategique mondial; vise à
Introduction générale - 1 - INTRODUCTION GENERALE I. Contexte et justification Le présent travail s’inscrit dans un double contexte: d’une part, un contexte stratégique mondial visant à réduire de moitié d'ici à 2015 le nombre de personnes n'ayant pas accès à l'eau potable et à l'assainissement, et ...
Page 26 - et
Introduction générale - 3 - La calibration du modèle consiste à rechercher les meilleures valeurs des paramètres cinétiques et stœchiométriques pour des conditions opératoires données, qui permettent d’obtenir les meilleures prédictions des valeurs mesurées de ces variables. Elle est généralement ré...
Page 27 - vise; . Il vient en complément à un modèle de connaissance relatif; faisant des stations; sur chacun de ses substrats potentiels,
Introduction générale - 4 - Le présent travail s’inscrit donc également dans ce contexte scientifique, et vise spécifiquement à améliorer la modélisation de la production du zooplancton dans les bassins de lagunage . Il vient en complément à un modèle de connaissance relatif aux bassins de lagunage ...
Page 28 - III. Plan de presentation du travail; a été testée
Introduction générale - 5 - III. Plan de presentation du travail Outre la présente introduction, ce travail se structure en huit chapitres. Le chapitre I fait un état général de la situation, vis-à-vis des objectifs de l’étude, et permet de mieux cerner les approches d’investigations mises en oeuvre...
Page 29 - Daphnia pulex
Introduction générale - 6 - Dans le chapitre VI, la stoechiométrie (les rapports de proportions dans lesquelles interviennent les substrats pour générer les différentes quantités de produits biomasses et métabolites) de chacun des processus de conversion biochimique retenus dans le sous-modèle est é...
Page 30 - on the population growth of
Introduction générale - 7 - Canovas S., Cassellas C., Picot B., Pena G. et Bontoux J. 1991. Evolution annuelle du peuplement zooplanctonique dans un lagunage à haut rendement et incidence du temps de séjour. Revue des sciences de l'eau / Journal of Water Science. 4 (2), 269-289. Cauchie H-M. 2000. P...
Page 31 - Chapitre I
Chapitre I - 9 - Chapitre I: ETAT DES CONNAISSANCES SUR LES SYSTEMES DE LAGUNAGE, LEUR ECOLOGIE, LA MODELISATION DE LA PRODUCTION DES CLADOCERES ET LES INTERACTIONS DES SUBSTRATS SUR LA CROISSANCE DES CLADOCÈRES I. Introduction Avant de développer ce chapitre, rappelons tout d’abord l’origine et le ...
Page 32 - II. Les differents systemes de lagunage et leur ecologie; « un bassin aménagé dans lequel l’eau usée est retenue pendant; le bassin de maturation
Chapitre I - 10 - Les procédés intensifs requièrent des coûts élevés en investissement et frais de fonctionnement ainsi qu’un personnel qualifié pour leur suivi et leur entretien (Pearson, 1995). Ils apparaissent à plusieurs égards très peu adaptés au contexte socio-économique des pays du sud. Par c...
Page 33 - si le lagunage naturel fonctionnait comme un réacteur
Chapitre I - 11 - Ouano (1981) distingue cinq types de bassins de lagunage : - Le bassin aérobie - Le bassin facultatif - Le bassin anaérobie - Le bassin de maturation - Le bassin aéré II.1.2 Principales caractéristiques Les différents types de bassins de lagunage se distinguent, les uns par rapport...
Page 34 - Schéma de fonctionnement d’un bassin facultatif (Tchobanoglous
Chapitre I - 12 - Figure I.1 : Schéma de fonctionnement d’un bassin facultatif (Tchobanoglous et al ., 1985), cités dans Shilton (2001) Dans la couche supérieure, l’oxygène apporté, d’une part par la diffusion à partir de l’atmosphère, mais surtout en présence de lu mière par la photosynthèse des al...
Page 35 - et CH
Chapitre I - 13 - II.1.2.2 Lagunage aérobie Les bassins aérobies se caractérisent essentiellement par leur faible profondeur 0,2 à 0,6 m qui assure une forte production algale et une bonne oxygénation. Ils permettent une élimination de 80 à 95% de pollution organique dissoute exprimée en DBO (Ouano,...
Page 36 - (2005) a relevé dans la littérature au moins; : Charge surfacique; kV; constante de dégradation; nQ; les
Chapitre I - 14 - Les stations d’épuration par le procédé du lagunage font généralement intervenir une ou plusieurs séries des bassins anaérobies, des bassins facultatifs suivis généralement des bassins de maturation en fonction de la qualité visée pour l’eau épurée (Varon et Mara, 2004). II.1.3 Dim...
Page 37 - charges organiques biologiques; dt; CV; QC
Chapitre I - 15 - La charge surfacique désigne la quantité de matières organiques, exprimées en Kg DBO 5 , appliquée à chaque hectare de la surface du bassin facultatif. Ouano (1981) a par ailleurs montré que les valeurs de références considérées pour la charge surfacique, varient selon les localité...
Page 38 - volumique
Chapitre I - 16 - II.1.3.1.3 Bassin anaérobies Ils sont préférentiellement dimensionnés sur la base de la charge organique volumique (Crites et al., 2006). D’une profondeur pouvant atteindre 7 m, ces bassins admettent des charges organiques volumiques variant selon les auteurs: 42 à 400g DBO 5 /(m 3...
Page 40 - al
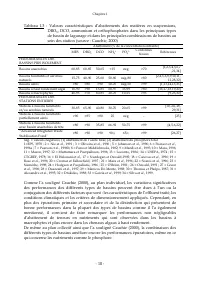
Chapitre I - 18 - Tableau I.3 : Valeurs caractéristiques d’abattements des matières en suspensions, DBO 5 , DCO, ammonium et orthophosphates dans les principaux types de bassin de lagunage et dans les principales combinaisons de bassins au sein des station (source : Cauchie, 2000) Abattement (% de l...
Page 41 - caractéristiques de l’eau usée; Volume d’eau
Chapitre I - 19 - Pour résumer, on peut retenir que le dimensionnement des bassins de lagunage suivant les modèles empiriques et rationnels se fait généralement sur la base d’une part, des caractéristiques de l’eau usée arrivant dans le bassin (en provenance de l’égout ou d’un bassin précédent), et ...
Page 43 - et al
Chapitre I - 21 - Bien calibré , le modèle ainsi obtenu, permet de réaliser (à partir de la résolution d’un ensemble d’équations différentielles) des simulations pour prédire l’évolution des variables du système ou pour servir à optimiser le dimensionnement d’ouvrages à partir des caractéristiques c...
Page 45 - II.2.1 Le zooplancton des bassins de lagunage et son écologie; ... l’ensemble des animaux
Chapitre I - 23 - o la consommation de l’H2 (S H2 ) par les méthanogènes hydrogénoclastes (X H2 ), o la consommation de l’H2 (S H2 ) par les B SR hydrogénoclastes (X H2 , So4) o l’oxydation des sulfures S H2S par les phototrophes sulfo-oxydantes X SOB . Chacun des processus de conversion biochimique...
Page 47 - Keratela sp; cyclopidés
Chapitre I - 25 - 1: Keratela sp ; 2: Cephalodella sp. ; 3: Trichocera sp. Figure I.4: Vue microscopique de quelques rotifères (Leclercq et Maquet, 2001) II.2.1.2.3 Les copépodes Ce sont des petits crustacés présents à la surface de l'eau, de forme généralement allongée ou vermififiée (figure I.5) s...
Page 49 - Les cladocères sont présents dans tous les bassins de lagunage
Chapitre I - 27 - Les cladocères sont présents dans tous les bassins de lagunage avec toutefois une répartition des taxons, dans les différents types de bassins de lagunage, répondant surtout au gradient d’oxydation de la matière. Les moinidae sont caractéristiques de biotopes hypereutrophes : Loedo...
Page 51 - par Mahapatra; III. Productions de zooplancton dans les bassins de lagunage
Chapitre I - 29 - Sous climat tropical, des espèces de flagellés appartenant aux genres des euglenophytes (Mahapatra et al. , 2013) ou des chlorophycées (Bernal et al. , 2008 ; cité par Mahapatra et al., 2013) semblent être les genres prédominants dans les conditions de fortes charges (bassins facul...
Page 52 - Daphnia sp; dessous par rapport à notre application, aux plans de leurs
Chapitre I - 30 - Tableau I.5 : Quelques ordres de grandeur de production et de taux de production Organisme Type de lagunage Production nette totale Référence bibliographique Non précisées Lagunage de Mèze 6000 g poids frais. m -2 .an -1 Argence cité par Barnabé (1983) Daphnia sp . Bassin pilote à ...
Page 53 - un sous-modèle mathématique
Chapitre I - 31 - IV.1 Le modèle de Hathaway et Stefan (1995) Afin de suivre l’évolution des populations de daphnies dans les bassins de lagunage du Minnesota dans la perspective de comprendre leur impact sur les biomasses algales, Hathaway et Stefan (1995) ont développé un sous-modèle mathématique ...
Page 54 - L’effet de la température sur la démographie des daphnies.; sous modèle; et d’un; sous-modèle biochimique; Il a été conçu en procédant à une sélection des
Chapitre I - 32 - - L’effet de la température sur la démographie des daphnies. La possibilité d’apprécier la qualité du modèle proposé vis-à-vis des objectifs pour lesquels il a été développé est mitigée par plusieurs facteurs: modèle calibré non testé, inadéquation des conditions dans lesquelles on...
Page 55 - La croissance; (Définition des sigles fournie en annexe); La mortalité est décrite par une cinétique de 1
Chapitre I - 33 - Trois processus portent sur le zooplancton: la croissance, la respiration et la mortalité. La croissance est décrite par une cinétique de type Monod corrigée conformément à Hirsch et Smale (1983) et Jacobsen (1983), cités par l’auteur, par un terme qui représente la croissance maxi...
Page 56 - IV.3 Le River Water Quality Model n°1 (Reichert
Chapitre I - 34 - Une tentative de traduire le modèle dans le formalisme de la matrice de Petersen en présentée en annexe I.2. Elle montre clairement l’absence de paramètres stœchiométriques et de coefficients stœchiométriques qui sont des éléments importants dans un modèle de conversion biochimique...
Page 59 - en poids sec par litre dans celui de Moreno-Grau
Chapitre I - 37 - - Les biomasses de daphnies ne sont pas exprimées dans la même unité dans ces différents modèles. Elles sont exprimées : o en nombre d’individus par litre dans le modèle de Hathaway et Stefen (1995), o en poids sec par litre dans celui de Moreno-Grau et al . (1996) et, o en poids p...
Page 60 - cyanophycées
Chapitre I - 38 - - Lorsqu’elles sont ingérées, les petites colonies et les fractions de formes filamenteuses pourraient être faiblement digérées ou assimilées ou pourraient manquer d’éléments essentiels à la croissance des cladocères ; - De nombreuses espèces et souches font preuves de toxicité env...
Page 61 - et que selon
Chapitre I - 39 - Dans sa revue de littérature, Melack (1985) a cité Saunders (1969) qui a montré que les daphnies assimilent très mal des particules détritiques stérilisées et que selon Rodina (1983), elles n’arrivent pas à se reproduire lorsqu’elles sont nourries par ces substrats. La qualité nutr...
Page 62 - production en; de; régulation; sus mentionnés. Les modèles recensés dans la littérature
Chapitre I - 40 - Par contre, l’analyse des modèles de lagunage ayant pris en compte le zooplancton couplée aux connaissances, d’une part sur l’écologie des bassins de lagunage et d’autre part, sur les effets des types de substrat sur la croissance des cladocères, révèle la nécessité d’approfondir l...
Page 63 - Moina
Chapitre I - 41 - Benider A., Pourriot R., Rougier C. et Tifnouti A. 1991. La population de Moina (Crustacea, cladocera) dans les bassins de lagunage de Marrakech (Maroc) : Moina macropa (Strauss, 1820) versus Moina micrura Kurtz, 1874. Rev. Hydrobiol. trop. 24 (3), 261-264. Buikema A. L. 1973. Filt...
Page 64 - cells by
Chapitre I - 42 - Hadas O., Bachrach V., Kott Y. et Cavari B.Z. 1983. Assimilation of E. coli cells by Daphnia magna on the whole organism level. Hydrobiologia. 102: 163-199. Halcrow K. 1976. The fine structure of the carapace integument of Daphnia magna Straus (Crustacea Branchiopoda). Cell. Tissue...
Page 65 - Webographie
Chapitre I - 44 - Ricard M., Stephan M., Loubet E, Bobillot J-P, Marie D., Saint-Dizier A. 1970. Atlas de biologie, Ed. Librairie Stock et Librairie Générale Française, traduit de la version allemande de Vogel G. et Angenmann H. 1967.Atlas zur Biologie. Deutscher Taschenbuch Verlag G.m .b.h. et Co, ...
Page 66 - Waste Stabilisation Ponds, IRC International Water and
Chapitre I - 45 - Khattabi H., 2002, Intérêts de l’étude des paramètres hydrogéologiques et hydrobiologiques pour la compréhension du fonctionnement de la station de traitement des lixiviats de la décharge d’ordures ménagères d’Etueffont (Belfort, France), Consulté le 15/07/2006, sur le site : http:...
Page 67 - Chapitre II; Hooper
Chapitre II - 46 - Chapitre II: COMPTAGE ET ESTIMATION DE BIOMASSE DE D. pulex PAR TRAITEMENT D’IMAGE I. Introduction Dans les conditions de laboratoire, deux principales démarches sont habituellement utilisées dans la littérature pour suivre les biomasses de zooplancton en général et de cladocères ...
Page 68 - II. Matériel et méthodes
Chapitre II - 47 - Le traitement d’image permet de suivre entièrement la même population durant toute la période d’une expérience en partant de la composition désirée d’individus (en termes de stade de développement). Il permet non seulement le comptage des individus mais également la mesure de leur...
Page 69 - Figure II.2: Deux photos successives du même lot de 115
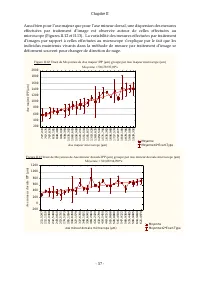
Chapitre II - 48 - II.2 Traitement d’image Après leurs captures, les images sont traitées conformément à Færovig et al. (2002), en réalisant une soustraction entre les deux images successives de chaque effectif (Figures II.2 à II.5). À l’aide du logiciel Image - Proplus (MediaCybernetics). La soustr...
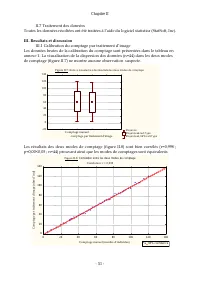
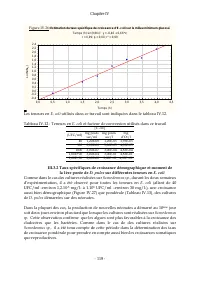
Page 72 - III. Resultats et discussion; III.1 Calibration du comptage par traitement d’image
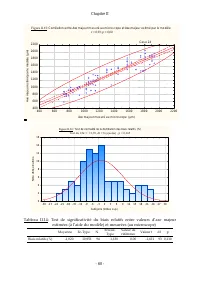
Chapitre II - 51 - II.7 Traitement des données Toutes les données récoltées ont été traitées à l’aide du logiciel statistica (StatSoft, Inc). III. Resultats et discussion III.1 Calibration du comptage par traitement d’image Les données brutes de la calibration du comptage sont présentées dans le tab...
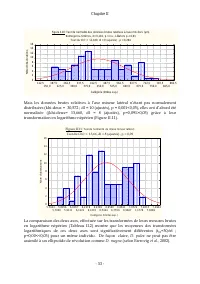
Page 74 - ne peut pas être
Chapitre II - 53 - Figure II.10: Tes t de normalité des données brutes relativ es à l'ax e min dors (µm), Kolmogorov -Smirnov , d= 0,110, p = n.s ., Lillief ors p < 0,01 Tes t du Chi² = 11,180, dl = 9 (ajustés) , p = 0,264 112,5 150,0 187,5 225,0 262,5 300,0 337,5 375,0 412,5 450,0 487,5 525,0 56...
Page 76 - Min Dors
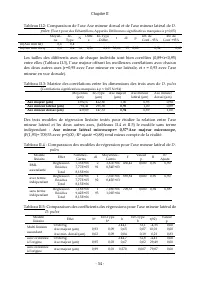
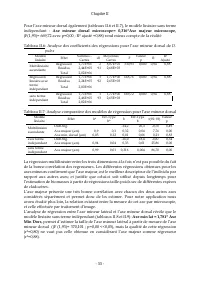
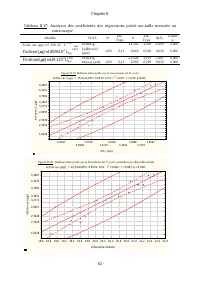
Chapitre II - 55 - Pour l’axe mineur dorsal également (tableaux II.6 et II.7), le modèle linéaire sans terme indépendant : Axe mineur dorsal microscope= 0,316*Axe majeur microscope, (F(1,93)= 669,72 avec p=0,00 ; R 2 ajusté =0,88) rend mieux compte de la réalité Tableau II.6: Analyse des coefficient...
Page 79 - reflète bien la taille réelle de l’axe majeur (r; le terme; Axe majeur par traitement d’image =; Analyse des coefficients des modèles d’estimation de taille par
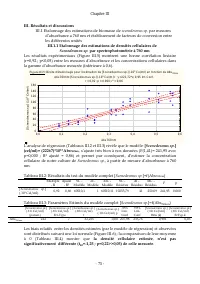
Chapitre II - 58 - Les résultats obtenus des analyses de régressions (Tableaux II.11 et II.12) révèlent que les régressions obtenues pour l’axe majeur sont meilleures (r 2ajusté ={0,85 ; 0,86} et F={537,103 ; 578,51} avec p<0,00 <<0,05 pour n=94) que celles obtenues pour l’axe mineur dorsal...
Page 80 - magna; un facteur
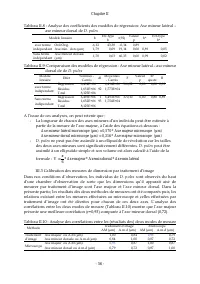
Chapitre II - 59 - Figure II. 14: Test de normalité de la distribution des biais relatifs des mesures d'axe majeur par traitement d'image par rapport aux mesures effectuées au microscope Kolmogorov-Smirnov, d= 0,114; p < 0, 20, Lilliefors p < 0, 01 T est du Chi ² = 6,72, dl = 3 (aj usté s) , p...
Page 81 - Axe m ajeur m es uré au m icros cope (μm )
Chapitre II - 60 - Figure II.15 : Corrélation entre Axe m ajeur m es uré au m icros cope et Axe m ajeur es tim é par le m odèle r = 0,93; p = 0,00 400 600 800 1000 1200 1400 1600 1800 2000 2200 Axe m ajeur m es uré au m icros cope (µm ) 400 600 800 1000 1200 1400 1600 1800 2000 2200 A x e m a je u r...
Page 82 - assimilés à des ellipsoïdes de révolution; mesurée au microscope; sec; Poids; Poids
Chapitre II - 61 - III.4 Relation poids sec - taille des individus La relation taille–poids sec est d’abord étudiée avec les tailles mesurées au microscope afin de pouvoir comparer nos résultats aux données de la littérature. Ensuite, les tailles sont converties en leurs valeurs mesurées par traitem...
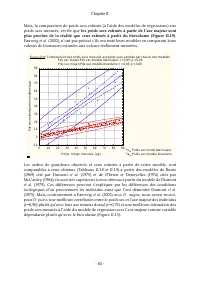
Page 84 - les poids secs estimés à partir de l’axe majeur sont; Færovig; ti; pour
Chapitre II - 63 - Mais, la comparaison de poids secs estimés (à l’aide des modèles de régressions) aux poids secs mesurés, révèle que les poids secs estimés à partir de l’axe majeur sont plus proches de la réalité que ceux estimés à partir du biovolume (Figure II.19) . Færovig et al. (2002), n’ont ...
Page 85 - majeur
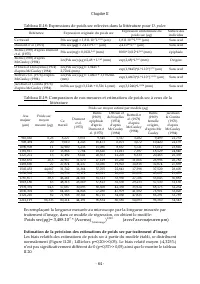
Chapitre II - 64 - Tableau II.18: Expressions de poids sec relevées dans la littérature pour D. pulex Référence Expression originale du poids sec Expression uniformisée du poids sec (µg) Nature des individus Ce travail Pds sec ( µg) = 1,811.10 -7 L 2,585 (µm) 1,811.10 -7 *L 2,585 (µm) Sans œuf Dumon...
Page 86 - Variable Moyenne Ec-Type
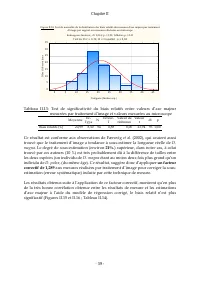
Chapitre II - 65 - Figure II.20 :T est de norm al i té d es bi ai s rel ati fs aux esti m ati ons de poi ds se c de D. pul ex pa r trai tem en t d 'i m age K-S d=0,089; p> 0,20; Lilliefors p> 0.20 -80 -6 0 -40 -20 0 20 40 6 0 X <= Borne de catégori e 0 1 2 3 4 5 N o m b re d 'o b s e rv a t...
Page 87 - Daphnia magna
Chapitre II - 66 - - Il s’avère donc possible d’appliquer le traitement d’image à l’estimation de l’abondance et de la biomasse de D. pulex , pour étudier la cinétique de sa croissance au laboratoire en estimant la biomasse (en poids sec) à l’aide de la formule : µm) en majeur axe l' (avec ) maj (Ax...
Page 88 - Chapitre III; Chapitre III : ÉTALONNAGE DES MESURES DE BIOMASSE DE SUBSTRAT; ont été définies à partir des données de la; densités cellulaires
Chapitre III - 67 - Chapitre III : ÉTALONNAGE DES MESURES DE BIOMASSE DE SUBSTRAT PAR SPECTROPHOTOMETRIE, ETUDES DE FACTEURS DE CONVERSION ENTRE UNITÉS ET APPLICABILITE DES BIOMOLES PROPOSEES DANS LE MODELE DE LAGUNAGE AUX SUBSTRATS ETUDIES I. Introduction En prélude aux études de cinétique et de st...
Page 89 - II. Matériels et Méthodes; Microcystis aeruginosa
Chapitre III - 68 - II. Matériels et Méthodes II.1 Origines des souches de substrat Scenedesmus sp. La souche de Scenedesmus sp. nous a été fournie par le Docteur Franck Fabrice du Laboratoire de Bioénergétique de l’université de Liège. Il s’agit d’une souche locale. M. aeruginosa La souche de Micro...
Page 90 - indiquent le sens de sélection des 20 cases comptées)
Chapitre III - 69 - II.3.1 Mesure de densité optique Les densités optiques ont été mesurées à l’aide d’un spectrophotomètre UV-3100PC (Figure III.1). Les mesures de densités optiques des différentes suspensions de substrats ( E. coli , M. aeruginosa et Scenedesmus sp .) ont été effectuées à des long...
Page 91 - dilution
Chapitre III - 70 - La concentration cellulaire se calcule à partir des comptages effectués à l’aide de cette cellule par la formule : dilution * 1000 * 0,1mm * 0,0025mm * 20) * (2 ) n + (n = l cellules.m de Nombre 2 2 1 1 Avec n 1 et n 2 , les nombres de cellules comptés respectivement dans 20 case...
Page 92 - filtré; Pour toutes les espèces (
Chapitre III - 71 - [ ] (ml) ensemencé volume dilution * colonie de Nombre (UFC/ml) coli E. = II.3.4 Estimations des matières en suspension (MES) et des Matières Volatiles en Suspension (MVS) Les estimations de MES (mg/ml), ont été déterminées conformément au protocole décrit par l’A.P.H. A. (1995)....
Page 94 - II.3.5 Mesures de DCO des biomasses de substrats; II.4 Etudes de facteurs de conversion entre différentes unités; Détermination indirecte; substrats étudiés
Chapitre III - 73 - II.3.5 Mesures de DCO des biomasses de substrats Les mesures de DCO ont été effectuées par la méthode photométrique à l’aide des tests en tubes DCO (Spectroquant, Merck). Une quantité suffisante de culture (suspension d’algue ou de bactérie) est d’abord complètement débarrassée d...
Page 95 - HPO; mole
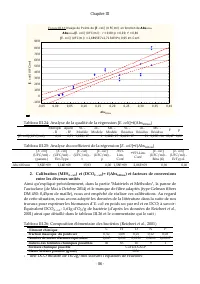
Chapitre III - 74 - A défaut de réaliser des analyses de composition élémentaire pour chacun des substrats considérés dans le présent travail, l’applicabilité des biomoles proposées dans le modèle de lagunage de l’unité "Eau et Assainissement" et dans le RWQM1 (Reichert et al., 2001) à ces s...
Page 96 - Scenedesmus sp
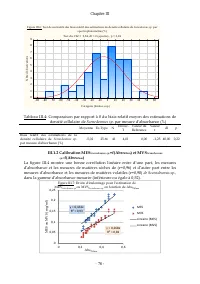
Chapitre III - 75 - III. Résultats et discussions III.1 Etalonnage des estimations de biomasse de scenedesmus sp . par mesures d’absorbance a 760 nm et établissement de facteurs de conversion entre les différentes unités III.1.1 Etalonnage des estimations de densités cellulaires de Scenedesmus sp. p...
Page 97 - densité cellulaire de; MES
Chapitre III - 76 - Figure III.6: Test de norma lité des biais rela tif des estima tions de densité cellulaire de Scenedesmus sp . pa r spectrophotométrie (%) Test du Chi² = 2 ,84, dl = 2 (a justés) , p = 0 ,24 -90 -80 -70 -60 -50 -40 -30 -20 -10 0 10 20 30 40 50 60 Catégorie (limites sup.) 0 1 2 3 ...
Page 98 - DCO
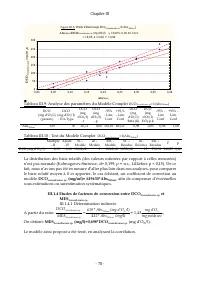
Chapitre III - 77 - Les analyses de régression (Tableaux III.5 et III.6) révèlent que le modèle: MES Scenedesmus sp. (mg/ml)= (0,432±0,006)*Abs 760nm , s’ajuste très bien à nos données (F(1,44)= 573,678 avec p<0,05 ; R 2 ajusté = 0,93) et permet par conséquent, d’estimer la teneur en poids sec de...
Page 99 - Tableau III.9: Analyse des paramètres du Modèle Complet; , afin de compenser d’éventuelles; III.1.4 Etudes de facteurs de conversion entre DCO
Chapitre III - 78 - Figure III.8: Droite d'étalonnage DCO Scenedesmus sp. =f (A bs 760nnm ) A bs 760nm : DCO Scenedesmus sp (mg d'O 2 /l): y = 619*x ; 0,95 Int. Conf . r = 0,95; p = 0,00; r² = 0,90 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 Abs 760nm 0 50 100 150 200 250 300 D C O S c e n e d e s ...
Page 100 - (Figure III.9), révèle que le modèle permet une bonne estimation des; Au; d’O; Mais, du fait que la distribution des biais relatifs n’est pas
Chapitre III - 79 - Figure III.9 : Analys e de corrélation entre MES es tim é via DCO es tim é et MES m es urées MES (mg/ml) :MES via DCO es tim é: r = 0,98; p = 0.00; r² = 0,97 MES via DCO es tim é = 0,0307+0,7958*x; 0,95 Int. Conf. 0,04 0,06 0,08 0,10 0,12 0,14 0,16 0,18 0,20 0,22 0,24 MES (mg/ml)...
Page 101 - de la biomole proposée pour; Elément chimique
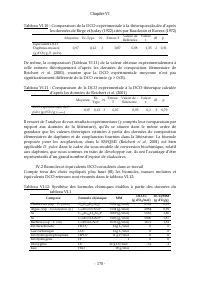
Chapitre III - 80 - Tableau III.11: Equivalents DCO (iDCO) déterminés expérimentalement MES (mg/l) DCO mesurée (mg d'O 2 /l) iDCO (mg d'O 2 /mg MES) MES (mg/l) DCO mesurée (mg d'O 2 /l) iDCO (mg d'O 2 /mg MES) 87,50 91 1,04 213,59 220 1,03 81,19 82 1,01 525,58 452 0,86 66,36 71 1,07 442,16 451 1,02 ...
Page 102 - Référence; III.1.6 Facteur de conversion entre densité cellulaire et MES pour
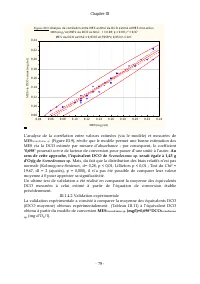
Chapitre III - 81 - Il ressort de cette équation que l’équivalent DCO théorique estimé à partir des données de Reichert et al . (2001) est de 0,95 g d’O 2 /g d’algue . La comparaison de la moyenne des mesures expérimentales de DCO de Scenedesmus sp ., (Tableau III.13) à l’équivalent DCO théorique dé...
Page 103 - us
Chapitre III - 82 - Le ratio entre les deux équations calibrations entre densité cellulaire et absorbance d’une part et, entre MES et absorbance d’autre part, permet d’estimer le poids sec d’une cellule de Scenedesmus sp ., à environ 1,95.10 -8 mg soit encore : 19,5 pg : cel / pg 5 , 19 = = 5 sp. s ...
Page 106 - Moyenne
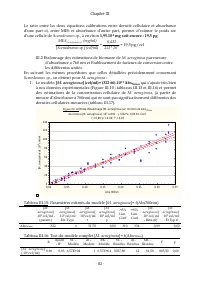
Chapitre III - 85 - De ce ratio, on peut établir: MES M. aeruginosa (mg/l)=1,059*DCO M. aeruginosa (mg d’O 2 /l). Les biais relatifs entre valeurs mesurées et valeurs estimées à l’aide de l’équation établie ci-dessus sont normalement distribués (Test du Chi² = 3,018 ; dl = 1 (ajustés), p= 0,08>0,...
Page 107 - ) et facteurs de conversions; en poids sec par ml et en DCO à savoir :; une DCO molaire de 1672g/mol suivant l’équation de réaction:
Chapitre III - 86 - Figure III.13: Nuage de Points de [E. coli] (UFC/m l) en fonction de Abs 600nm Abs 600nm :[E. coli] (UFC/m l) : r = 0,89; p = 0,00; r² = 0,80 [ E. coli] (UFC/m l) = 2,6895E7+1,7134E9*x; 0,95 Int. Conf. -0,05 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 Abs 600nm -1E8 0 1E8 2E8 3E...
Page 110 - Chapitre IV; Chapitre IV: CARACTERISATION DE LA CINETIQUE DE CROISSANCE DE; SUR
Chapitre IV - 89 - Chapitre IV: CARACTERISATION DE LA CINETIQUE DE CROISSANCE DE D. pulex SUR Scenedesmus sp ., E. coli , et M. aeruginosa I. Introduction En biotechnologie, la cinétique de croissance (production) d’un organisme, décrit la vitesse à laquelle cet organisme est produit dans un réacteu...
Page 111 - II. Matériels et méthodes; II.1 Justification du plan expérimental; de croissance; Exp
Chapitre IV - 90 - Une cinétique de type inhibition de croissance s’observe lorsque le substrat est favorable à la croissance de l’organisme étudié avec toutefois, une diminution du taux de croissance en présence des fortes concentrations de substrat. La cinétique de type mortalité conviendrait s’il...
Page 113 - additive
Chapitre IV - 92 - Somme de cinétiques de croissance avec paramètre d’interaction Ce type de modèle ( proposé par Yoon et al ., 1977, cité par Reardon et al. , 2000) est utilisé pour décrire des types d’inhibitions non spécifiques entre deux ou plusieurs substrats homologues , lorsque les interactio...
Page 114 - les formes multiplicatives ne sont pas adaptées pour les; II.1.2 Détermination des paramètres cinétiques
Chapitre IV - 93 - Pour Reardon et al. (2000), les formes multiplicatives ne sont pas adaptées pour les substrats homologues (dans notre cas : E. coli , Scenedesmus sp . et M. aeruginosa ), car si la teneur d’un substrat s’approchait de zéro, la fonction de croissance tendrait aussi vers zéro, alors...
Page 115 - plusieurs substrats (potentiels) vivants
Chapitre IV - 94 - II.1.2.2 Adaptation de la méthode aux daphnies Ainsi que le soulignait Reardon et al. (2000), malgré que la croissance des microorganismes sur des substrats mixtes est rencontrée dans la nature, la caractérisation mathématique de la cinétique de leur croissance sur ces substrats m...
Page 116 - Configuration générale du système de culture
Chapitre IV - 95 - Avec la découverte de milieux synthétiques susceptibles de supporter à la fois la croissance des algues et des daphnies tels que le milieux Combo (Kilham et al ., 1998), il est devenu encore plus facile de mettre en œuvre des systèmes de cultures en continu qui supportent à la foi...
Page 118 - ln; rt
Chapitre IV - 97 - II.1.2.4 Détermination du taux de croissance de D. pulex sur chaque concentration en substrat Dans les conditions favorables de culture, la croissance démographique des daphnies est considérée comme exponentielle et, généralement décrite par le modèle de Krebs (1985), cité par Alv...
Page 119 - ml; Scendesmus sp
Chapitre IV - 98 - - Le taux de croissance maximale dans nos conditions expérimentales, de chacune de ces deux espèces de substrat cultivée en chémostat soit respectivement 0,73 j -1 pour Scenesdesmus sp . et 0,50 j -1 Microcystis aeruginosa ; - La concentration finale visée de substrat (C f. substr...
Page 120 - Cf; Le milieu combo (Kilham
Chapitre IV - 99 - En définitive, les calculs de gestion du chémostat ont été effectués à l’aide du tableur Excel. Un exemple est fourni des variables et paramètres considérés dans les calculs est fourni dans le tableau IV.2. Du fait de la relation qui lie la concentration finale (Cf) visée dans les...
Page 122 - respiration endogène
Chapitre IV - 101 - Après chaque estimation de biomasse (chaque deux jours), les cuves de culture de daphnie sont remplacées par d’autres plus propres. De même, le contenu de chaque cuve est remplacé par le volume et la concentration en substrat requis. II.3 Estimation et suivi des biomasses Toutes ...
Page 123 - Temps; déterminés graphiquement; Alors que l’évolution du logarithme de la densité de
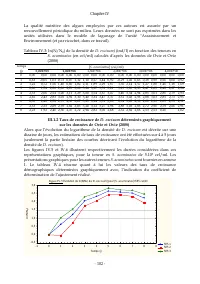
Chapitre IV - 102 - La qualité nutritive des algues employées par ces auteurs est assurée par un renouvellement périodique du milieu. Leurs données ne sont pas exprimées dans les unités utilisées dans le modèle de lagunage de l’unité "Assainissement et Environnement (et par ricochet, dans ce tra...
Page 124 - III.1.3 Caractérisation de la cinétique de croissance de
Chapitre IV - 103 - Figure IV.6 :Taux de crois s ance (r) de D. Excisum pour [S. acum inatus]=5E5 cel/m l Tem ps (j):5E5 1: y = 0,1104 + 0,55*x; r = 0,99; p = 0,00; r² = 0,97 Tem ps (j):5E5 2: y = -0,51 + 0,63*x; r = 0,98; p = 0,00; r² = 0,95 Tem ps (j):5E5 3: y = 0,32 + 0,56*x; r = 0,97; p = 0,00; ...
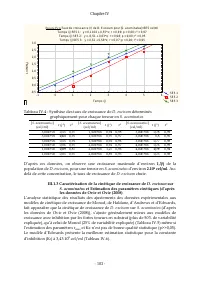
Page 125 - Monod
Chapitre IV - 104 - Tableau IV. 5 : Comparaison des ajustements des taux de croissance de D. excisum à divers modèles de cinétique de croissance Modèle Effet Somme des carrés DL Moindres carrés Valeur F Valeur p r variance expliquée (%) Monod Régression 12,45 2 6,22 287,19 0,00 0,50 25 Résidu 0,35 1...
Page 127 - paramètres; Conclusion partielle; sur
Chapitre IV - 106 - III.1.5 Vérification sommaire de la fiabilité de ces estimations des paramètres L’analyse sommaire de la fiabilité des estimations (Tableau IV.7 et Figure IV.11) révèle que le modèle d’Edwards décrit effectivement mieux la cinétique de la croissance de D. excisum sur S. acuminatu...
Page 128 - sur le milieu combo; concentrations cellulaires au départ; de Scenedesmus sp
Chapitre IV - 107 - Tableau IV.8 : Les teneurs Scenedesmus sp considérées pour l’étude de la cinétique de croissance de D. pulex [ Scenedesmus sp .] Cel/ml mg/l mg d’O 2 /l 9,42E+03 0,18 0,17 2,22E+05 4,32 4,10 1,11E+06 21,60 20,5 2,22E+06 43,20 41,04 4,04E+06 78,55 74,62 1,21E+07 235,64 223,86 III....
Page 129 - détermination du moment de la 1ère ponte; Pour toutes les teneurs en; pulex sur les différentes teneurs en Scendesmus sp.; jour de
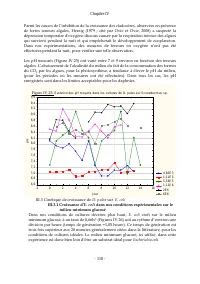
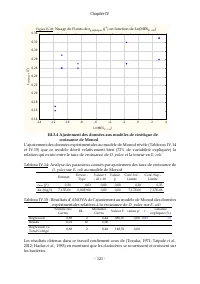
Chapitre IV - 108 - III.2.2 Évolution des taux spécifiques de croissance démographique et détermination du moment de la 1ère ponte Pour toutes les teneurs en Scenedesmus sp. utilisées dans ces expérimentations (de 0,18 mg/l (soit 9,42.10 3 cel/ml) à 235,64mg/ml (1,21.10 7 cel/ml)), il a été observé ...
Page 130 - III.2.3 Taux spécifiques de croissance pondérale de; Coefficient de; Coefficient de
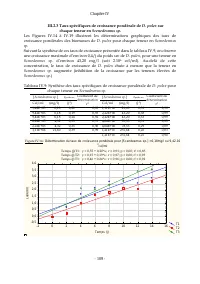
Chapitre IV - 109 - III.2.3 Taux spécifiques de croissance pondérale de D. pulex sur chaque teneur en Scenedesmus sp. Les Figures IV.14 à IV.19 illustrent les déterminations graphiques des taux de croissance pondérales des biomasses de D. pulex pour chaque teneur en Scenedesmus sp . Suivant la synth...
Page 132 - : Détermination du taux de croissance pondérale pour [
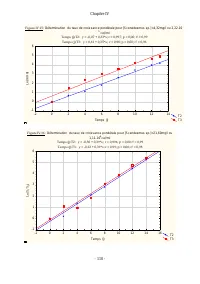
Chapitre IV - 111 - Figure IV.17 : Détermination du taux de croissance pondérale pour [ Scenedesmus sp. ]=43,20mg /l ou 2,22.10 6 cel/ml Temps (j):T2: y = -0,07 + 0,38*x; r = 0,99; p = 0,00; r² = 0,99 Temps (j):T3: y = 0,07 + 0,33*x; r = 0,996; p = 0,00; r² = 0,99 T2 T3 -2 0 2 4 6 8 10 12 14 Temps (...
Page 133 - III.2.4 Ajustement des données expérimentales aux modèles de; montre que tous les modèles
Chapitre IV - 112 - Figure IV.19 : Déterm ination du taux de crois s ance pondérale pour [Scenedesm us sp.]=235,64m g/l ou 1,21.10 7 cel/m l Temps (j):T1: y = 0,61 + 0,26*x; r = 0,91; p = 0,00; r² = 0,83 Temps (j):T2: y = -0,21 + 0,25*x; r = 0,95; p = 0,00; r² = 0,90 T1 T2 -2 0 2 4 6 8 10 12 14 16 T...
Page 134 - mg
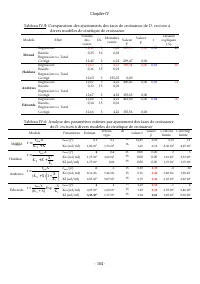
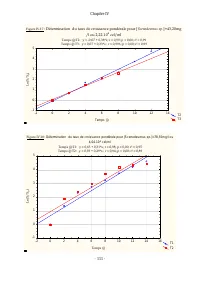
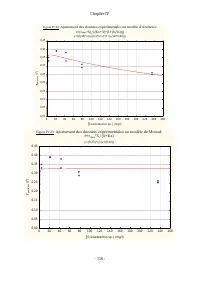
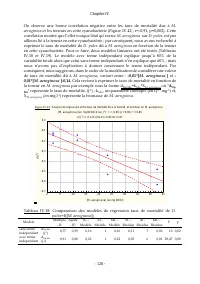
Chapitre IV - 113 - Tableau IV.10: Comparaison des ajustements des données expérimentales relatives à la croissance de D. pulex sur Scenedesmus sp. à divers modèles de cinétique de croissance Modèle Effet Somme des carrés DL Moindres carrés Valeur F Valeur p r % de la variance expliquée Monod Régres...
Page 135 - des paramètres estim
Chapitre IV - 114 - Les estimations des valeurs des paramètres cinétiques dans notre cas, pour la croissance de D. pulex sur Scenedesmus sp . sont de meilleures qualités statistiques que celles obtenues en traitant les données de Ovie et Ovie (2008) concernant la croissance de D. excisum sur S. acum...
Page 136 - : Ajustement des données expérimentales au modèle d'Edwards:; : Ajustement des données expérimentales au modèle de Haldane:
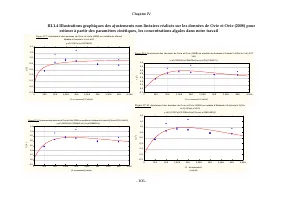
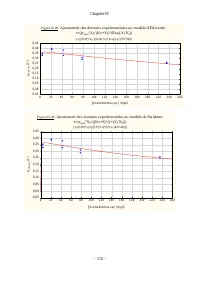
Chapitre IV - 115 - Figure IV.20 : Ajustement des données expérimentales au modèle d'Edwards: r=((r max *X)/(Kx+X))*(Exp(-X/K I )) y=(((0,37)*x)/((0,14)+x))*(exp(-x/(577,36))) 0 20 40 60 80 100 120 140 160 180 200 220 240 260 [ Scenedesm us sp.] (m g/l) -0,05 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 ...
Page 137 - : Ajustement des données expérimentales au modèle de Monod:
Chapitre IV - 116 - Figure IV.22 : Ajustement des données expérimentales au modèle d'Andrews: r=(r max *X)/((Kx+X)*(1+(X/K I ))) y=((0,38)*x)/(((0,15)+x)*(1+(x/(459,43)))) 0 20 40 60 80 100 120 140 160 180 200 220 240 260 [ Scenedesmus sp.] (mg/l) 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 0,45 r ...
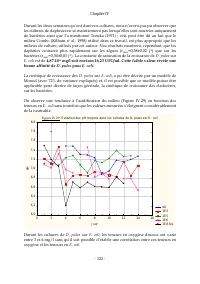
Page 138 - III.2.5 Suivi des variables physico-chimiques lors des cultures de; : Evolution des moyennes de teneurs en oxygène
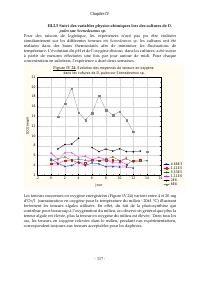
Chapitre IV - 117 - III.2.5 Suivi des variables physico-chimiques lors des cultures de D. pulex sur Scenedesmus sp . Pour des raisons de logistique, les expériences n’ont pas pu être réalisées simultanément sur les différentes teneurs en Scenedesmus sp. les cultures ont été réalisées dans des bains ...
Page 139 - Jour; III.3 Cinétique de croissance de; dans nos conditions expérimentales sur le; Dans nos conditions de cultures décrites plus haut,
Chapitre IV - 118 - Parmi les causes de l’inhibition de la croissance des cladocères, observées en présence de fortes teneurs algales, Herzig (1979 ; cité par Ovie et Ovie, 2008) a suspecté la dépression temporaire d'oxygène dissous causée par la respiration intense des algues qui survient pendant l...
Page 140 - mg poids; III.3.2 Taux spécifiques de croissance démographique et moment de
Chapitre IV - 119 - Figure IV.26 : Estimation du taux spécifique de croissance d'E. coli sur le milieu minimum glucosé Tem ps (h):Ln(N/N 0 ): y = -0,46 + 0,66*x; r = 0,99; p = 0,00; r² = 0,99 0,0 0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0 4,5 Tem ps (h) -0,4 -0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2...
Page 142 - : Nuage de Points de; III.3.4 Ajustement des données aux modèles de cinétique de
Chapitre IV - 121 - Figure IV.28 : Nuage de Points de r graphique (j -1 ) en fonction de Ln(MES E. coli ) -14 -12 -10 -8 -6 -4 -2 0 2 4 Ln(MES E. c oli ) 0,14 0,16 0,18 0,20 0,22 0,24 0,26 0,28 0,30 0,32 0,34 r g ra p h iq u e ( j -1 ) III.3.4 Ajustement des données aux modèles de cinétique de crois...
Page 143 - bonne affinité de; : Evolution des pH moyens dans les cultures de D. pulex sur E. coli
Chapitre IV - 122 - Durant les deux semaines qu’ont duré nos cultures, nous n’avons pas pu observer que les cultures de daphnies ne se maintiennent pas lorsqu’elles sont nourries uniquement de bactéries ainsi que l’a mentionné Tezuka (1971) ; cela peut être dû au fait que le milieu Combo (Kilham et ...
Page 144 - : Droite d'estimation du taux spécifique de croissance de
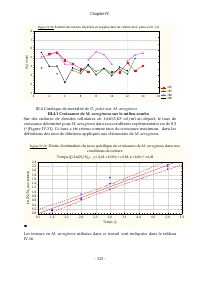
Chapitre IV - 123 - Figure IV.30 : Evolution des teneurs moy ennes en ox ygène dans les cultures de D. pul ex s ur E. col i 1E2 1E5 1E6 1E8 0 2 4 6 8 10 12 14 16 1 2 3 4 5 6 7 8 [O 2 ] (m g /l ) III.4 Cinétique de mortalité de D. pulex sur M. aeruginosa III.4.1 Croissance de M. aeruginosa sur le mil...
Page 145 - Équivalent DCO; III.4.2 Évolution des taux spécifiques de mortalité de; Figure IV.32: Evoluti on des taux spécifiques de mortalité de; ne peut pas être considéré comme substrat
Chapitre IV - 124 - Tableau IV.16: Teneurs en M. aeruginosa utilisées dans ce travail [ M. aeruginosa ] Cel/ml mg/l Équivalent DCO (mg d'O 2 /l) 2,00E+06 44,91 42,42 1,10E+05 2,47 2,33 1,10E+06 24,70 23,33 5,50E+05 12,35 11,67 III.4.2 Évolution des taux spécifiques de mortalité de D. pulex sur M. ae...
Page 146 - face à; Ln
Chapitre IV - 125 - La détermination des taux spécifiques de mortalité en poids/poids.jour pour chacune des teneurs en M. aeruginosa est relativement plus difficile que ceux exprimés en nombre d’individus/nombre d’individus.j (Figures IV.34 à IV.41). Mais face à l’évidence de la mortalité , nous avo...
Page 147 - LN
Chapitre IV - 126 - -1,50 -1,00 -0,50 0,00 0,50 1,00 1,50 2,00 0 2 4 6 8 10 LN ( X /X 0 ) Temps (jour) Figure IV.34: Evolution Ln(X/X 0 ) pour [ M. aeruginosa ]=2E6 cel/ml T1 T2 y = -0,4833x + 3,8691 R² = 0,8176 y = -0,627x + 4,1611 R² = 0,8829 -1,50 -1,00 -0,50 0,00 0,50 1,00 1,50 2,00 2,50 0 5 10 ...
Page 150 - préfère être seule plutôt qu’en
Chapitre IV - 129 - Tableau IV.19: Analyse des coefficients des modèles taux de mortalité de D. pulex =f([ M. aeruginosa ]) Modèle d Dp-Ma (j-1) (param.) d Dp-Ma (j -1 ) Err-Type d Dp-Ma (j -1 ) t d Dp-Ma (j -1 ) p -95% Lim Conf +95% Lim. Conf d Dp-Ma (j -1 ) Bêta (ß) d Dp-Ma (j -1 ) - ErTyp.ß -95% ...
Page 152 - III.6.1 Taux de croissance pondérale sur différentes mixtures de; III.6.2 Caractérisation de la cinétique globale
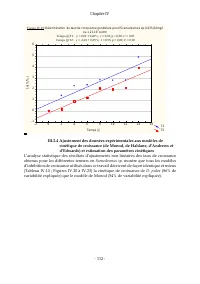
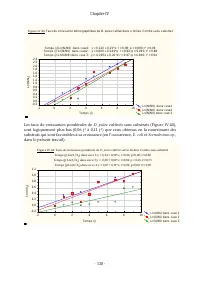
Chapitre IV - 131 - III.6 Cinétique globale de croissance de D. pulex sur ses substrats III.6.1 Taux de croissance pondérale sur différentes mixtures de Scenedesmus sp . et E. coli Au regard des résultats obtenus des études réalisées en considérant les substrats individuellement, la cinétique global...
Page 153 - III.6.3 Estimation des paramètres d’interaction
Chapitre IV - 132 - Avec les définitions des symboles conformes à celles données dans la partie "Matériels et Méthodes" III.6.3 Estimation des paramètres d’interaction Nos données expérimentales s’ajustent très bien au modèle de cinétique de croissance avec paramètres d’interaction (≈80% de ...
Page 154 - substrats hétérologues
Chapitre IV - 133 - Afin de tester la pertinence de considérer l’interaction ainsi relevé entre Scenedesmus sp. et E. coli , sur la croissance de D. pulex , nous avons comparé les ajustements de nos données, d’une part au modèle de cinétique de croissance avec paramètre d’interaction, à ceux réalisé...
Page 156 - Chapitre V; Chapitre V: DESCRIPTION CONCEPTUELLE DU SOUS - MODELE; , de matières organiques particulaires; processus de croissance; ). Certains de ces produits contribuent d’ailleurs à stimuler la
Chapitre V - 137 - Chapitre V: DESCRIPTION CONCEPTUELLE DU SOUS - MODELE "PRODUCTION DES CLADOCERES DANS LES BASSINS DE LAGUNAGE" I. Introduction Au regard des résultats obtenus de la caractérisation de la cinétique de croissance des cladocères (chapitre IV) qui montrent clairement que seule...
Page 157 - . La vitesse de ce processus qui occasionne; processus de mortalité dû à la présence des cyanobactéries
Chapitre V - 138 - En considérant une biomôle constante pour toutes les classes de taille de daphnies, le sous-modèle décrit une relation stœchiométrique entre la biomasse de daphnie et celles des substrats qu’elle consomme d’une part, et les produits qu’elle excrète d’autre part, au cours de ce pro...
Page 158 - respiration endogène (k; mortalité naturelle (k; mortalité due aux cyanobactéries (k; QX
Chapitre V - 139 - Figure V.1: Description schématique du modèle proposé Synthèse Dans le sous-modèle ici proposé, le taux net de croissance des daphnies dans un bassin de lagunage, résulte du solde entre les taux respectifs des processus de croissance (r croissance ) , de respiration endogène (k re...
Page 161 - substrats; l’accepteur final d’électron
Chapitre V - 142 - II.1.3 Processus de mortalité (avec ou sans l’influence des cyanobactéries) Ce processus traduit la perte de biomasse active de cladocères (par effet de vieillissement) sans que cela soit associé à une consommation d’oxygène. La biomasse active ainsi perdue va accroitre la matière...
Page 164 - stœchiométries
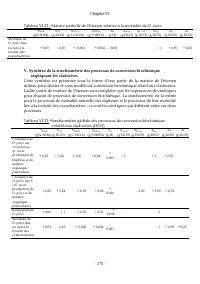
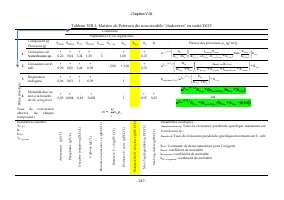
Chapitre V - 145 - Leur biomasse est exprimée en équivalent DCO par litre ou en poids sec par litre. III. Expression du sous modèle dans le formalisme de matrice des processus Le formalisme de matrice des processus (Henze et al. , 2000) est un format de description synthétique des modèles de convers...
Page 165 - matrice de Petersen; d’élaboration pour le sous-modèle relatif aux cladocères
Chapitre V - 146 - Le formalisme de matrice des processus (Henze et al. , 2000) facilite aussi la vérification de la conservation de la matière relativement à chaque élément chimique dans chaque processus, soit : ∑ 0 = t ij α υ . Parfois, seule la matrice des stœchiométries est présentée, accompagné...
Page 166 - il; ou; ri
Chapitre V - 147 - Continuité Substances et/ou organismes B il an m as si q u e Composé (i) Processus (j) S NH4 S HPO4 S O2 S HCO3 X Scene X E. coli X D. p X M.a X S X I Vitesse des processus, ρ j (g/l.j) 1 Croissance sur algues + + - + - +1 + p D. Scene I Scene coli E. . coli/Scene E. Scene X Scene...
Page 168 - Symbole
Chapitre V - 149 - Tableau V.2: Les paramètres cinétiques Symbole Définition Unité r max,D.p , Scenedesmus sp , T° Taux de croissance spécifique maximum de D. pulex sur Scenedesmus sp. j -1 r max,D.p , E. coli , T° Taux de croissance spécifique maximum de D. pulex sur E. coli. j -1 Kx, algue Constan...
Page 170 - Par contre, le processus de mortalité a été reformulé pour
Chapitre VI - 152 - Cet effet toxique létal qu’exercent les cyanobactéries sur les cladocères doit être pris en compte pour bien modéliser la production de cladocères dans tout système . A notre connaissance, aucun autre modèle n’avait abordé cet aspect de la modélisation. Omlin et al. (2001) ont bi...
Page 171 - Chapitre VI; une; équivalent DCO théorique de 2984g d’O
Chapitre VI - 153 - o La période d’acclimatation, avant la mesure du taux d’ingestion, (Buikema, 1973) o La densité des cladocères dans le milieu (Helgen, 1987) o La température, dont l’effet varie en fonction de la concentration en substrat (Kersting, 1978). La composition élémentaire en C, N et P ...
Page 172 - III. Matériels et méthodes; coefficients stœchiométriques
Chapitre VI - 154 - III. Matériels et méthodes Dans le présent travail, les deux approches de comptabilité de la matière utilisées dans le RWQM1 (Reichert et al ., 2001) ont été adoptées pour décrire la stœchiométrie de chacun des quatre processus de conversion biochimique dans lesquels interviennen...
Page 176 - quantité (en g de poids
Chapitre VI - 158 - De même que pour le rendement de production de daphnies, le concept de taux d’excrétion utilisé ici (en modélisation) diffère de celui généralement employé en hydrobiologie. Dans ce travail, on s’intéresse pour une même période, à la quantité (en g de poids sec) de substrat excré...
Page 177 - vitesse d’accumulation; Rhodotorula glutinus
Chapitre VI - 159 - h 1 * ) daphnie sec poids g ( α * daphnie) sec (poids g * h substrat) de sec (poids g = ) substrat de sec poids ( mX ∆ substrat Les lignes qui suivent présentent les méthodes pour déterminer le taux d’ingestion et la croissance pondérale. o Mesure du taux d’ingestion Peters (1984...
Page 178 - mesurer l’activité de la culture mère
Chapitre VI - 160 - La justification donnée à ce choix est que même s’il a été rapporté que la durée minimale du transit intestinal de D. pulex est supérieure à 10 min (Pacaud, 1939), l’égestion post abdominale du substrat marqué pourrait démarrer pour des fortes concentrations de substrat, après en...
Page 179 - Il s’avère évident par conséquent que l’on ne saurait en; quantité de substrat excrétée
Chapitre VI - 161 - 5 ème étape: Calcul du taux d’ingestion Les taux d’ingestion seront déterminés sur chacun des substrats: Scenedesmus sp . et E. coli. Mesure de la variation pondérale des cladocères "∆m XDaphnie " La croissance somatique des cladocères survient de manière périodique et pr...
Page 180 - La durée de l’; leur présentation dans le formalisme de la matrice de Petersen
Chapitre VI - 162 - La terminologie anglaise distingue "Egestion" qui correspond à la fraction des aliments non assimilés à travers le tube digestif et éliminés sous forme de fèces et, "excretion" qui correspond aux déchets résultants de l’assimilation. Dans le présent travail de mod...
Page 181 - fe; DCOmolaire; les quantités de réactifs et de produits
Chapitre VI - 163 - Substrat i D.p i X. - X X D a X D = consommé substrat molaire DCO a produite biomasse molaire DCO = ingéré substrat DCO g produite biomasse DCO g = Y S D.p molaire CO molaire CO alors, ubstrat S D.p X p D. X D Y D a molaire CO molaire CO × = (1) o Calcul du coefficient stœchiomét...
Page 182 - les quantités (en DCO) des réactifs et des produits, par; II.3.1 Expression du processus de respiration
Chapitre VI - 164 - II.2.3.2.1 Cas des composés à DCO non nuls Les coefficients stœchiométriques précédemment calculés sur base de relations molaires entre les réactifs et les produits, sont présentés dans le formalisme de Pertersen en calculant les quantités (en DCO) des réactifs et des produits, p...
Page 184 - II.4.2 Valeurs des paramètres stœchiométriques du processus de; : traduit la fraction de; a été maintenue pour ce paramètre stœchiométrique; Le Taux de minéralisation; originaire des cladocères, qui devient inerte suite à leur mort:
Chapitre VI - 166 - Dans cette démarche, il est proposé que s’il n’y a pas une forte évidence de différence de compositions élémentaires pour les différentes classes de matières organiques, l’on utilise la même composition élémentaire pour les algues, les consommateurs (donc ici, les cladocères) et ...
Page 185 - processus de mortalité; de la matrice de Petersen
Chapitre VI - 167 - II.4.3 Détermination des valeurs des coefficients stœchiométriques du processus de mortalité Les valeurs des huit (8) coefficients stœchiométriques inconnus (a, b, c, d, e, f, g et h) sont déterminées en résolvant un système d’équation comprenant : - Les deux (2) équations relati...
Page 186 - IV.1.2 Estimation d’une biomole théorique et de la DCO
Chapitre VI - 168 - Tableau VI.4: Analyse de l’influence de la température de séchage sur la valeur de la fraction organique (Test t pour des Echantillons Indépendants) Moyenne - Groupe 1 Moyenne - Groupe 2 valeur t dl p N Actifs Groupe 1 N Actifs Groupe 2 Ec-Type Groupe 1 Ec-Type Groupe 2 Ratio F -...
Page 187 - IV.1.3 Mesures expérimentales réalisées sur nos cultures de; Deux mesures ont été effectuées sur nos cultures de; Ce travail; littérature
Chapitre VI - 169 - IV.1.3 Mesures expérimentales réalisées sur nos cultures de D. pulex Deux mesures ont été effectuées sur nos cultures de D. pulex . Il d’agit d’une part, de mesures de DCO, et d’autre part, de composition élémentaire. Leurs résultats sont présentés respectivement dans les tableau...
Page 188 - applicable; Composé
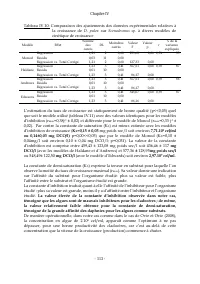
Chapitre VI - 170 - Tableau VI.10 : Comparaison de la DCO expérimentale à la théoriquecalculée d’après les données de Birge et Juday (1922) cités par Baudouin et Ravera (1972) Moyenne Ec-Type N Erreur-T Valeur de Référence Valeur t dl p Equivalent DCO Daphnies mesuré (g d’O 2 /g D. pulex ) 0,97 0,12...
Page 189 - dans mon texte
Chapitre VI - 171 - IV.3 Stœchiométrie du processus de croissance IV.3.1 Croissance sur Scenedesmus sp. Le processus de la croissance de D. pulex sur Scenedesmus sp. est conceptuellement décrit par l’équation de réaction é.2. En considérant les valeurs des paramètres stœchiométriques indiquées, dans...
Page 190 - avons
Chapitre VI - 172 - A cette étape de la description du processus de croissance de D. pulex , on observe que pour produire 1g d’équivalent DCO de D. pulex, 5 g d’équivalent DCO de Scenedesmus sp., sont oxydés dont 0,76 sont convertis en matières organiques particulaire et, le reste (soit 3,24 équival...
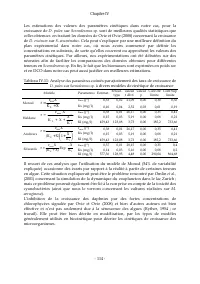
Page 192 - processus de mortalité de; l’équation de réaction traduisant le processus de respiration; Gauche; (2001) est lié au fait d’utiliser des biomoles
Chapitre VI - 174 - La matrice de Petersen relative au processus de respiration de D. pulex , peut alors être présentée partiellement (Tableau VI.19). Tableau VI.19: Matrice partielle de Petersen relative à la respiration de D. pulex S NH4 (gN-NH4) S O2 (g DCO) S HCO3 (g C-HCO 3 ) S HPO4 (g P-HPO 4 ...
Page 193 - impliquant les cladocères
Chapitre VI - 175 - Tableau VI.22 : Matrice partielle de Petersen relative à la mortalité de D. pulex S NH4 (gN-NH4) S O2 (g DCO) S HCO3 (g C-HCO 3 ) S HPO4 (g P-HPO 4 ) S H (g H) X Scene (g DCO) X E. Coli (g DCO) X Dp (g DCO) Xs (g DCO) Xi (g DCO) Mortalité de D. pulex due ou non à la toxicité des ...
Page 194 - . Cependant, nous avons effectué des commentaires
Chapitre VI - 176 - VI. Conclusion La comparaison de nos données expérimentales sur la composition élémentaire de D. pulex , aux données de la littérature y compris celles considérées dans le RWQM1 (Reichert et al., 2001), a montré que la biomole proposée pour le zooplancton dans le RWQM1 (Reichert ...
Page 196 - Chapitre VII; Chapitre VII : ANALYSE DE RENTABILITE D’UN AVANT-PROJET DE; I.1 Contexte et justification; manque de rentabilité financière des projets; nombreuses expériences de valorisation du plancton; (produit dans des bassins; en pisciculture; ont été capitalisées depuis trois décennies à travers le; valorisation indirecte; du zooplancton en pisciculture : elles
Chapitre VII - 180 - Chapitre VII : ANALYSE DE RENTABILITE D’UN AVANT-PROJET DE VALORISATION DES CLADOCERES PRODUITS DANS LES BASSINS DE LAGUNAGE AU BENIN I. Introduction I.1 Contexte et justification Le contexte du présent projet, développé en détail dans l’introduction générale de la thèse, se car...
Page 197 - valorisation directe du phytoplancton; pour produire des; valorisation indirecte du
Chapitre VII - 181 - - Les expériences de valorisation directe du phytoplancton pour produire des poissons phytoplanctonophages correspondent aux travaux, plus récents, d’Edwards (1992 cité par Mara et al. , 1993); et de Mara et al. (1993). Leur principe pourrait être appliqué pour produire des espè...
Page 198 - II. Description de l’avant- projet et des données de base
Chapitre VII - 182 - I.2 Objectifs de l’avant-projet I.2.1 Objectif général Contribuer aux efforts de facilitation de l’accès à l’assainissement pour tous dans les pays du sud, en intégrant des incitants financiers dans les projets d’assainissement. I.2.2 Objectif spécifique Estimer la rentabilité f...
Page 199 - Bassin de maturation
Chapitre VII - 183 - En effet d’après Cowey (1979) cité par Guérrin (1988) la teneur en protéine du zooplancton déshydraté suffit à couvrir les besoins quantitatifs de la plupart des poissons étudiés ; ces besoins sont compris entre 38% et 46% de l’aliment sec, pour la carpe commune, la truite arc-e...
Page 200 - II.2.1 Dimensions des bassins et de l’exploitation; La concentration en polluant (DBO
Chapitre VII - 184 - L’autre raison qui justifie, dans notre contexte, la préférence pour les bassins de maturation est leur faible exigence en terres (1 ha environ) comparé aux 6 ha qui auraient été nécessaires dans notre contexte (§C.2) pour le bassin à poisson. Ainsi, les caractéristiques des bas...
Page 201 - la surface moyenne (S; anaérobie est de 500m; La charge volumique (L
Chapitre VII - 185 - Les caractéristiques moyennes des eaux usées, d’un pays donné, sont généralement exprimées sous la forme d’équivalent-habitant c’est-à-dire la quantité de pollution générée par un habitant. Leur connaissance combinée à la connaissance de l’effectif de la population ciblée par le...
Page 202 - ts
Chapitre VII - 186 - Pour une profondeur (p fac ) de 1,5m, le débit (Q) de 1000m 3 /j, et un temps de séjour (ts fac ) de 5 jours proposé par Mara (1993), on obtient une surface moyenne (S fac ) à la mi- profondeur du bassin facultatifs, de 3333m 2 (cal cul ci-dessous): 2 1 3 fac fac fac m 3333 = m ...
Page 203 - Ce; : respectivement, les temps de rétention hydraulique (temps
Chapitre VII - 187 - Ce : la concentration en coliforme dans l’effluent (UFC/100 ml) ts ana , ts fac , ts mat,1 et ts mat,2 : respectivement, les temps de rétention hydraulique (temps de séjour) dans le bassin anaérobie, dans le bassin facultatif et dans le bassin de maturation. k : Constante d’abat...
Page 205 - II.2.2 Production journalière de biomasse
Chapitre VII - 189 - II.2.1.6 Le Dessableur Le dessableur permet de retenir les particules denses dont la vitesse de chute est comprise entre 0,2 et 0,3 m/s (principalement le sable) en amont de la station pour éviter qu’ils s’accumulent dans les bassins, réduisant leur volume utile pour l’eau usée,...
Page 208 - Dépenses de fonctionnement
Chapitre VII - 192 - III. Méthode La méthode proposée pour analyser la viabilité économique du projet, repose sur une analyse de filière pour chacune de ses deux variantes, et une comparaison de leur rentabilité financière. III.1 Analyse de filière L’analyse de filière, a été faite suivant une démar...
Page 209 - récoltée; IV. Résultats et discussions
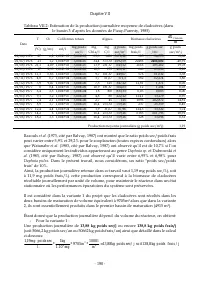
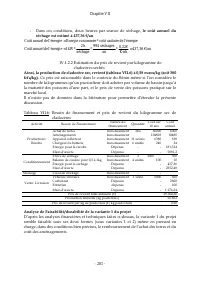
Chapitre VII - 193 - III.2 Analyse de rentabilité financière La rentabilité financière de chaque variante est analysée en comparant le coût de revient au prix de vente possible, pour chacune unité du produit généré (daphnies fraiches ou sèches dans le cas de la variante 1, ou poisson dans le cas de ...
Page 210 - Achat de terres; décomposé comme suit :; Achat d’appareils collecteurs
Chapitre VII - 194 - IV.1.1.2 Identification et estimation des besoins de financement IV.1.1.2.1 Identification des besoins de financement par activité Tableau VII.3: Besoins de financements identifiés pour la sous variante 1 du projet Activité Besoin de Financement Nature du financement Production ...
Page 211 - Cap
Chapitre VII - 195 - o Une batterie qui assure la fourniture d’énergie o Un contacteur o Un coffre de protection étanche équipé de deux poignets de portage - Le dispositif de pêche. Il comprend : o Un flotteur qui assure la flottation du système en maintenant le moteur émergé o Un moteur (12V/20W) q...
Page 212 - Achat de chargeur de batteries; Données de base
Chapitre VII - 196 - Tableau VII.4 : Estimation détaillée du prix de revient d’un appareil- collecteur Rubrique Composante Montant (€) Energie Batterie 12V, 110Ah 180 Coffre étanche de protection de la batterie 108 Câble de rechargement pour batterie auto 25 Poignets de portage du coffre 20 Support ...
Page 213 - Puissance du chargeur (P; Durée de chargement (T; Besoins en sac de congélation
Chapitre VII - 197 - Coût d’un rechargement d’une batterie - Caractéristiques du chargeur de la batterie : o Tension (U chargeur ):12V o Intensité (I chargeur ):12A o Puissance du chargeur (P chargeur ) : 12V *12A =144W soit 0,144kW - Caractéristiques de la batterie : o Durée de chargement (T charge...
Page 214 - Balance; Nombre de congélateur nécessaire
Chapitre VII - 198 - an an semaines semaine j rouleau j rouleaux / € 6 , 1068 52 * 5 * € 37 , 1 * 3 = Balance Il est prévu d’acquérir quatre balances de cuisine, pour un coût total de 100 € pour les pesées. Main d’œuvre pour le conditionnement Il est prévu l’équivalent de deux personnes pour une rém...
Page 215 - Véhicule de livraison; Le coût annuel de carburant est estimé à; Main d’œuvre pour la commercialisation; daphnies fraiches
Chapitre VII - 199 - IV.1.1.2.2.4 Activités de commercialisation Véhicule de livraison Un véhicule utilitaire d’occasion est proposé pour un coût estimatif de 5000€ Coût du carburant Le coût annuel de carburant est estimé à 2600€/an an an semaines semaine l l / € 2600 52 * 50 * € 1 = Un forfait annu...
Page 216 - IV.1.2 Sous variante 2 : Production et vente de cladocères séchés; IV.1.2.1 Identification et estimation des besoins de financement
Chapitre VII - 200 - Ce prix de revient peut être ramené à 0,27 €/kg de poids frais, si l’achat des terres et leur aménagement ne sont pas pris en charge par le projet. IV.1.1.4 Estimation du prix de vente par kilogramme de daphnies fraiches En admettant un prix de revient de 0,60 €/kg (si le rembou...
Page 217 - Dans ces conditions, deux heures par séance de séchage,; le coût annuel du; cladocères séchés
Chapitre VII - 201 - - Dans ces conditions, deux heures par séance de séchage, le coût annuel du séchage est estimé à 437,36 €/an . énergie l' de unitaire coût * conommée Energie énergie l' de annuel Coût = €/an 36 , 437 Kwh 0,22€ * an séchages 994 * séchage h 2 * kW 1 énergie l' de annuel Coût = = ...
Page 218 - mm
Chapitre VII - 202 - Les bénéfices susceptibles d’être réalisés sont considérables dans l’hypothèse d’un bénéfice correspondant à la moitié du prix de vente fixé (14.752 €/ha.an à 15.083 €/ha.an) si l’on considère la surface totale de l’exploitation (et non uniquement celle des bassins de maturation...
Page 219 - IV.2.2 Estimation de la biomasse maximale de Alestes baremoze; Alestes baremoze; baremoze
Chapitre VII - 203 - Au-delà, on trouve des coefficients d'allométrie nettement supérieurs à 3 et différents chez les mâles et les femelles. La croissance pondérale doit donc être calculée en fonction de ces différents paramètres et surtout en tenant compte des variations saisonnières de condition q...
Page 220 - (1993) ont seulement considéré la surface des deux bassins de
Chapitre VII - 204 - ( ) l'élevage de durée initial) (Poids final) (Poids TCS ln ln − = Dans le cas présent, en considérant un seul individu, on obtiendra un taux TCS de 0,32/mois comme détaillé dans le calcul ci-dessous: ( ) 1 1 0,01.j ou 32. , 0 mois 20 ln(0,14g) ln(90,06g) TCS − − = − = mois Ce t...
Page 221 - IV.2.3 Description de la filière de valorisation proposée
Chapitre VII - 205 - Dans le cadre d’une gestion pérenne de cette activité, 5 poissons devraient être admis à poursuivre leur développement jusqu’à la production du nombre d’alevins requis pour le prochain cycle de production. Pour rappel, une femelle 220 g porte en moyenne 51.000 œufs (Durand, 1978...
Page 222 - Main d‘œuvre; pendant 2 heures, il faudrait; l’équivalent de 38 personnes; le coût total de la main d’œuvre; ainsi que détaillé dans l’équation ci-dessous :; Achat de congélateur
Chapitre VII - 206 - IV.2.4.2.2 Activités de récolte Achat de matériels Tableau VII.9 : Estimations des besoins de financement pour la récolte des poissons Matériel Prix unitaire (€) Quantité Montant (€) Grande senne 381,68 20 7633,6 Epuisette 7,63 20 152,6 Bassine 7,63 20 152,6 Peson 3,82 20 76,4 V...
Page 223 - Congélateur
Chapitre VII - 207 - En considérant que le conditionnement durera au maximum un mois pendant chaque cycle de production de 20 mois, d’après ces données de base , le coût annuel de l’énergie nécessaire pour le stockage annuel est de 6,22 €/an estimé conformément au calcul présenté ci-dessous : mois r...
Page 224 - IV.2.6 Estimation du prix de vente du kilogramme de poissons; (1993) et qui porte sur une production de
Chapitre VII - 208 - Tableau VII.11: Besoins de financement et prix de revient du poisson Activité Besoin de financement Nature du financement Cout sur dix ans Coût annuel Production Achat de terre Investissement 60000 6000 Aménagement Investissement 104803 10480 Achat d’alevins Investissement 6037,...
Page 228 - Chapitre VIII; Chapitre VIII: EBAUCHE POUR L’ETUDE APPRONFONDIE DE LA; pour répondre à la question:; Est-ce que le modèle proposé dans la présente
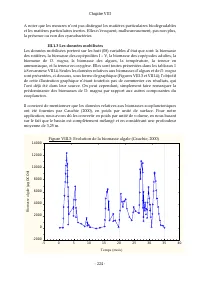
Chapitre VIII - 212 - Chapitre VIII: EBAUCHE POUR L’ETUDE APPRONFONDIE DE LA COHERENCE DU MODELE ET DE L’IMPACT DES CLADOCERES SUR L’EPURATION DES EAUX DANS LES BASSINS DE LAGUNAGE I. Introduction Après avoir mis en exergue les insuffisances relevées dans les modèles de la littérature, pour décrire ...
Page 229 - quel est le; qu’ils permettent de réaliser. L’impact des; II.1 Analyse de cohérence
Chapitre VIII - 213 - Plus clairement, nonobstant l’effet des saisons (qui gouvernent par la température, par l’insolation, et par d’autres facteurs, une dynamique des populations aquatiques), en considérant des caractéristiques constantes pour l’eau usée entrant dans un bassin de lagunage, l’on dev...
Page 230 - , les matières en suspension, les biomasses algales; le type de; II.4 Sources des données expérimentales
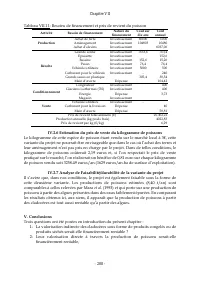
Chapitre VIII - 214 - II.2 Analyse de l’impact des cladocères sur les performances épuratoires L’impact de la présence des cladocères sur les performances épuratoires est analysé en comparant, à l’état stationnaire, un bassin contenant des cladocères et un bassin témoin (ne contenant pas de cladocèr...
Page 231 - grille
Chapitre VIII - 215 - Chacune des sources de données exploitées a été présentée suivant une grille comportant les quatre principaux points que sont : - Les caractéristiques des bassins : type de bassin de lagunage, charge organique appliquée, temps de séjour, dimensions du bassin, type d’hydrodynami...
Page 235 - étape : Définition des modèles mathématiques; étape : Construction de la configuration graphique; en pratique; une phase de sédiments.
Chapitre VIII - 219 - II.7 Simulation dans WEST Comme tout logiciel de simulation, WEST permet de résoudre les multiples équations différentielles inhérentes aux systèmes biologiques dans lesquels chaque modèle traduit un processus spécifique. Le logiciel WEST présente entre autres avantages, de com...
Page 238 - étape : Définition et réalisation des tests de simulation
Chapitre VIII - 222 - - Teneurs en azote total - Teneurs phosphates. A ces variables, celles relatives aux biomasses d’algues, de bactéries, de cyanobactéries, et de cladocères peuvent été ajoutées, dans la mesure du possible. 5 ème étape : Définition et réalisation des tests de simulation La simula...
Page 239 - in situ
Chapitre VIII - 223 - III.1. Les données de Cauchie (2000) Le travail de Cauchie (2000) a porté sur le suivi de la dynamique du métazooplancton et de la production secondaire de Daphnia magna dans un bassin de lagunage aéré (artificiellement). III.1.1 Caractéristiques des bassins Les caractéristique...
Page 240 - io
Chapitre VIII - 224 - A noter que les mesures n’ont pas distingué les matières particulaires biodégradables et les matières particulaires inertes. Elles n’évoquent, malheureusement, pas non plus, la présence ou non des cyanobactéries. III.1.3 Les données mobilisées Les données mobilisées portent sur...
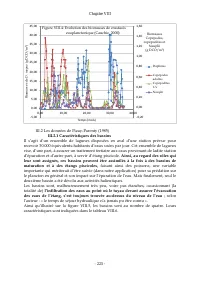
Page 241 - Ainsi, au regard des rôles qui; faisant ainsi des poissons, une variable; Biomasses; Figure VIII.4: Evolution des biomasses de crustacés
Chapitre VIII - 225 - III.2 Les données de Pizay-Parenty (1985) III.2.1 Caractéristiques des bassins Il s’agit d’un ensemble de lagunes disposées en aval d’une station prévue pour recevoir 30.000 équivalents-habitants d’eaux usées par jour. Cet ensemble de lagunes vise, d’une part, à assurer un trai...
Page 243 - Seul le débit à l’entrée de la serpentine a pu être estimé :; initialement prévu pour; III.2.2 Vérification des fonctions des bassins
Chapitre VIII - 227 - Seul le débit à l’entrée de la serpentine a pu être estimé : initialement prévu pour 400 m 3 /j à 500 m 3 /j, il a dû souvent être monté à 1200 m 3 /j ou 1500 m 3 /j pour compenser les pertes par infiltration. Conséquemment, la charge organique surfacique appliquée sur le bassi...
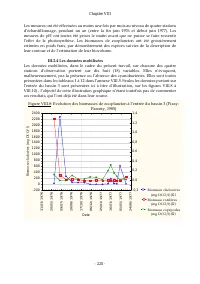
Page 244 - Biomasse cladocères
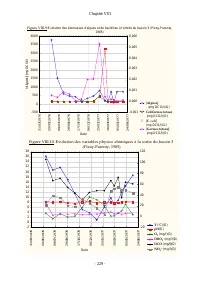
Chapitre VIII - 228 - Les mesures ont été effectuées au moins une fois par mois au niveau de quatre stations d’échantillonnage, pendant un an (entre la fin juin 1976 et début juin 1977). Les mesures de pH ont toutes été prises le matin avant que ne puisse se faire ressentir l’effet de la photosynthè...
Page 245 - Coliformes totaux; lg; Date
Chapitre VIII - 229 - Figure VIII.9 :Evolution des biomasses d'algues et de bactéries à l'entrée du bassin 3 (Pizay-Parenty, 1985) [ Algues] ( mg DCO/l)(G) Coliformes totaux ( mg DCO/l)(D) [ E. coli ] (mg DCO/l)(D) [ Germes totaux] ( mg DCO/l)(D) 3 1 /0 3 /1 9 7 6 2 0 /0 5 /1 9 7 6 0 9 /0 7 /1 9 7 6...
Page 246 - copie photographiée; . Ces données ont été; III.3.2 Vérification des fonctions des bassins; soit; DBO
Chapitre VIII - 230 - III.3 les données de Moreno-Clavel et al. (1990) cités par Moreno-Grau et al. (1996) Les données expérimentales de Moreno-Clavel et al . (1990) cités par Moreno-Grau et al. (1996) et les travaux y relatifs ont été publiés en espagnol par Moreno-Clavel et al . (1990) dont, nous ...
Page 249 - III.4.2 Vérification des fonctions des bassins
Chapitre VIII - 233 - A partir des valeurs initiales des variables, et des expressions mathématiques employées pour les diverses cinétiques, les auteurs ont montré des résultats de simulation portant sur : - Les bactéries en suspension - Le phytoplancton (facteur de conversion proposé par Jørgensen ...
Page 250 - . Cette charge est conforme au critère de Mara et Pearson (1986)
Chapitre VIII - 234 - Tableau VIII.11: Estimation des charges appliquées et vérification des fonctions des bassins (Source: Kawai et al. 1987) Paramètre Effluent Eau usée brute Bassin anaérobie 1 er Bassin facultatif Effluent 2 ème Bassin facultatif Bassin à Daphnies DBO (mg/l) 284 118 61,6 32,5 13,...
Page 251 - bassin facultatif
Chapitre VIII - 235 - Tableau VIII.12: Les données mobilisées dans Kawai et al. (1987) Paramètre Eau usée brute Effluent Bassin anaérobie 1 er Bassin facultatif 2 ème Bassin facultatif Bassins à daphnies DBO (mg/l) 284 118 61,6 32,5 13,2 DCO (mg/l) 534 234 196,3 125,4 118,3 N kjeldahl (mg/l) 15,77 1...
Page 252 - III.5.2 Vérification de la fonction du bassin considéré pour la; ha
Chapitre VIII - 236 - Les caractéristiques du bassin facultatif auquel l’ensemble des bassins est assimilé sont indiquées dans le tableau VIII.13. Tableau VIII.13: Caractéristiques du bassin facultatif considéré par Sunarsih et al. (2013) Paramètre Valeur Unité Volume de bassin 43680 m 3 Temps de sé...
Page 255 - cycle, Université des Sciences et Techniques de Lille, 195 p.
Chapitre VIII - 239 - Guérrin F. 1988. Valorisation du zooplancton produit en étangs de lagunage comme base pour l'alimentation de larves et juvéniles de cyprinidés. Bull. Fr. Pêche Piscic. 311: 113-125. Harerimana C. 2011. Modélisation de l'activité des bactéries du soufre en lagunage Thèse de Doct...
Page 256 - Conclusions – Recommandations - Perspectives; CONCLUSIONS – RECOMMANDATIONS - PERSPECTIVES
Conclusions – Recommandations - Perspectives - 241 - CONCLUSIONS – RECOMMANDATIONS - PERSPECTIVES Conclusions La présente thèse s’est intéressée à l’amélioration de la modélisation du fonctionnement des bassins de lagunage à travers la prise en compte des processus de conversions biochimiques impliq...
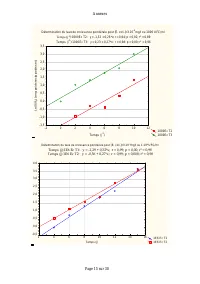
Page 257 - le sous-modèle est; Pour produire 1g d’équivalent DCO de
Conclusions – Recommandations - Perspectives - 242 - Ces résultats constituent une importante contribution à la modélisation du fonctionnement de bassins de lagunage du fait qu’ils spécifient clairement les modèles qui décrivent les cinétiques de croissance des cladocères sur leurs substrats et la c...
Page 258 - variante 1 consistant en une vente des
Conclusions – Recommandations - Perspectives - 243 - En attendant, les analyses cinétiques et stœchiométriques montrent que dans des conditions non limitantes en substrats et en absence de cyanobactéries, la vitesse de croissance des cladocères étant supérieure à la somme des vitesses de leur respir...
Page 260 - Références
Conclusions – Recommandations - Perspectives - 245 - Le contexte stratégique mondial qui vise à promouvoir l’accès à l’assainissement constitue une véritable opportunité, à travers les financements qui sont mis à disposition par la communauté internationale, pour les projets d’assainissement. L’exis...
Page 261 - Annexes; Densité
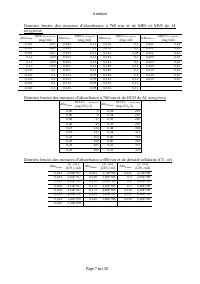
Annexes Page 1 sur 30 Annexes Annexe I.1 : Essai de présentation du sous- modèle de Hathaway et Stefan (1995) dans une matrice de Petersen Processus O 2 NH 3 H 2 S X Algue X Daphnie Xs Taux de croissance (mg/l.j) 1. Reproduction parthénogénétique - - + ue lg A ue lg A X ue lg A max X + K X r * X Dap...
Page 262 - Processus; Ss
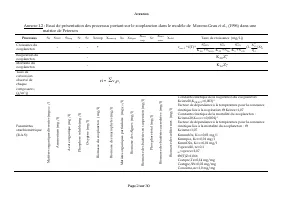
Annexes Page 2 sur 30 Annexe I.2 : Essai de présentation des processus portant sur le zooplancton dans le modèle de Moreno-Grau et al ., (1996) dans une matrice de Petersen Processus Ss S NH3 S Norg S P S O Xzoop X macrop Xs X Algue X bact susp X P X bact accro X colif. Taux de croissance (mg/l.j) C...
Page 271 - Evolution du taux spécifiques de croissance démographique
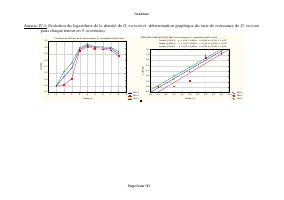
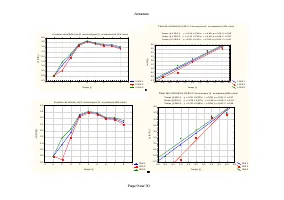
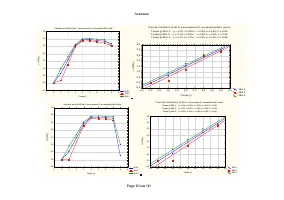
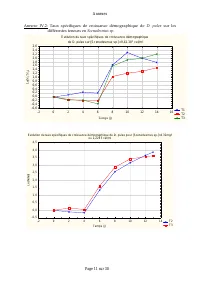
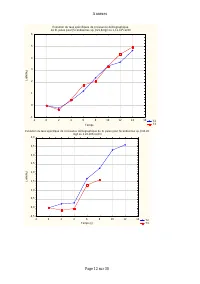
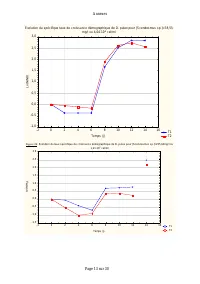
Annexes Page 11 sur 30 Annexe IV.2: Taux spécifiques de croissance démographique de D. pulex sur les différentes teneurs en Scenedesmus sp. Evolution du taux spécifiques de croissance démographique de D. pulex sur [Scenedesmus sp.]=9,42.10 3 cel/ml T1 T2 T3 -2 0 2 4 6 8 10 12 14 16 Temps (j) -0,6 -0...
Page 278 - QUESTIONNAIRE
Annexes Page 18 sur 30 Annexe VIII.2 : QUESTIONNAIRE 1- Combien de personnes et combien de temps faut-il pour dans un bassin de 500m 2 ? 2- De quels moyens doivent-ils disposer ? - - - - - - 3- Pour chaque matériel cité, donner une estimation du prix d’achat et de la quantité requise Matériel Estima...
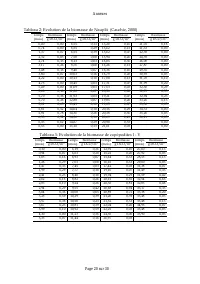
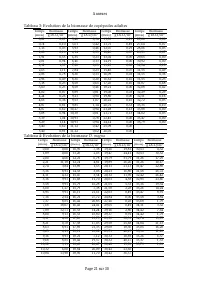
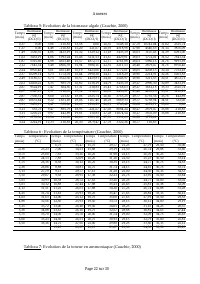
Page 279 - Tableau 1: Evolution de la biomasse de rotifères
Annexes Page 19 sur 30 Annexe VIII.3: Les valeurs des paramètres utilisés pour estimer la production journalière de cladocères Symbole Définition Valeur Unité r max,D.p , Scenedesmus sp , T° Taux de croissance spécifique maximum de D. pulex sur Scenedesmus sp. 0,37± 0,01 j -1 r max,D.p , E. coli , T...